Потенциал действия, его фазы. Современное представление о механизме его генерации
Восстанавливается за счёт диффузии из клетки положительно заряженных ионов калия, концентрация которых в окружающей среде также значительно ниже внутриклеточной.
Энциклопедичный YouTube
1 / 5
Электротонические потенциалы и потенциалы действия
Потенциал действия кардиомиоцитов
Потенциал действия рабочего миокарда
Потенциал действия в клетках-пейсмейкерах
Мембранные потенциалы - Часть 1
Субтитры
В предыдущем видеоролике речь шла о том, как клетка использует натрий-калиевый насос и АТФ для поддержания разницы потенциалов между внутренним пространством клетки и внешней средой. В целом, внешняя среда более положительно заряжена, чем внутренняя. У нас имеется разница потенциалов -70 милливольт между внутренней и внешней средой. Это значение со знаком минус, поскольку внешняя среда более положительно заряжена. Если из менее положительного значения вычесть более положительное значение, то у вас получится отрицательное значение порядка -70. Это утверждение является основой для понимания того, каким образом нейроны передают сигналы в организме. И чтобы лучше объяснить это, я хочу дать вам еще одно понятие дополнительно. После этого вы поймете, в чем состоит принцип передачи сигнала нейроном в реальной жизни. И более того, вам станет понятно, для чего им необходимы эти миелиновые оболочки и перехваты Ранвье, и для чего нужны все эти дендриты. Я надеюсь, у вас сложится целостная картина. Итак, есть два пути, которыми может перемещаться потенциал. Два пути передачи сигнала. Назовем это так.. Я не знаю какое слово подходит больше. Итак, первый путь - электротонический. Звучит очень сложно, но вы убедитесь, что в основе лежит очень простая идея. Сначала запишу сам термин. Электротонический... электротонический потенциал. И второе, на чем я собираюсь остановиться, это потенциал действия. У обоих из них имеются положительные и отрицательные стороны в контексте способности передачи сигнала. Мы будем говорить о них в рамках представлений о клетке и клеточной мембране. Давайте теперь со всем этим подробно разберемся. Итак, я изображу мембрану клетки. Условимся, что это нервная клетка, или нейрон, ведь мы обсуждаем именно нервные клетки. Мы знаем, что более положительный заряд находится на внешней стороне.. Мы также знаем, что на внешней стороне имеется большое количество натрия, и Его количество на внешней стороне больше, чем на внутренней стороне. Здесь его может быть совсем немного. И мы знаем, что внутри содержание калия намного больше, чем снаружи, однако мы также знаем, что внешняя сторона имеет больший положительный заряд, чем внутренняя, поскольку наш натрий-калиевый насос будет выкачивать три иона натрия на каждые два иона калия, которые он закачивает внутрь. И в последнем видеоролике я говорил вам, что существуют структуры, которые называются натриевыми воротами. Ворота для ионов натрия. Вот это все ионы. Они обладают зарядом. И теперь допустим, что имеются некоторые обстоятельства, некоторые стимулы - давайте я отмечу их. Опущусь немного ниже. Вот здесь расположены наши ворота для ионов натрия. Они находятся в закрытом положении, однако давайте скажем, что нечто вызывает их открытие. Мы поговорим, возможно, в этом видеоролике, или в этом и следующем видеороликах о различных вещах, которые могут вызывать их открытие. Определенный тип стимула вызывает их открытие. На самом деле, существует целый ряд различных стимулов. В общем, допустим, они открылись. Что же происходит вслед за их открытием? Мы открыли их, точнее это сделал определенный стимул - что же произойдет дальше? У нас имеется более положительный заряд на внешней стороне, чем на внутренней стороне, поэтому положительно заряженные объекты захотят проникнуть внутрь. А это натриевые ворота, поэтому только натрий может проникать через них. Это, своего рода, изогнутая белковая структура, через которую могут проникать только ионы натрия. И кроме всего прочего, у нас намного больше натрия снаружи, чем внутри. Поэтому градиент диффузии будет способствовать тому, что натрий будет проходить по нему. А в результате того, что ионы натрия заряжены положительно, и внешняя среда также более положительно заряжена, они будут стремиться покинуть эту положительно заряженную среду. Итак, если вы откроете эти ворота, то у вас будет большое количество ионов натрия, готовых к проникновению. Нарисую тут их побольше. И теперь, что должно произойти, если мы будем двигаться дальше по мембране? Давайте увеличим изображение. Итак, давайте представим, что наша мембрана. А это наши открытые ворота, они по какой-то причине открылись, и большое количество ионов натрия проникает внутрь. И все это становится более положительно заряженным. Давайте представим, что здесь у нас есть вольтметр. Мы измеряем разницу потенциалов между внутренней поверхностью мембраны и внешней поверхностью. Давайте я изображу небольшую схему. Я нарисую схему вот здесь на моем вольтметре. И это будет разница потенциалов - или мы можем назвать ее мембранным вольтажем или разницей вольтажа между двумя поверхностями мембраны - и давайте обозначим эту ось как время. Представим, что мы еще не открывали эти ворота. То есть это состояние покоя. Наши натрий-калиевые насосы продолжают работать. Частицы просачиваются назад и вперед, однако соотношение удерживается на значении минус 70 милливольт. Итак, вот здесь у нас минус 70 милливольт. И теперь, как только эти ворота, которые располагаются в некоторых других отделах клетки, открываются, что же происходит? Допустим, при этом, что открываются только эти ворота. Итак, здесь внезапно образуется более положительный заряд. Итак, положительные заряды, здесь уже есть - поэтому другие положительные заряды, обусловленные ионами натрия или калия, будут стремиться быть как можно дальше от этой точки, потому что в этой зоне отсутствует поток положительных ионов. То есть, она менее положительно заряжена, по сравнению с вот этим участком. У нас тут имеется некоторое количество ионов калия, и возможно, некоторое количество ионов натрия. Все они будут стремиться быть как можно дальше от того места, где открылись ворота. Заряд будет стремиться распространиться как можно дальше. И как только это происходит, как только мы открываем эти ворота, у нас происходит смещение положительного заряда в этом направлении. Это происходило при минус 70 милливольтах. Итак, некоторое количество положительного заряда уходит. Все это происходит очень быстро. Практически немедленно после этого, заряд становится менее отрицательным, или более положительным. Разница потенциалов между этим и этим участками становится меньше. Это соответствует данной точке на графике. И теперь, если мы берем эту точку, если мы проделаем то же самое - если мы измерим вольтаж в этой точке вот здесь, то возможно, он будет минус 70 милливольт, а некоторое время спустя, положительный заряд начнет оказывать свое влияние и заряд здесь станет более положительным, при этом эффект будет уменьшаться. Поскольку эти положительные заряды будут распространяться во всех направлениях. В результате эффект как бы разрежается. То есть он становится менее выраженным. И заряд здесь станет менее положительным. Итак, электротонический потенциал происходит в одной точке клетки, когда ворота открываются и заряд начинает распространяться внутрь, и начинает воздействовать на потенциал в других отделах клетки. Однако положительным моментом является то, что он очень быстрый. По мере того, как это происходит... По мере поступления в клетку, он становится все более и более положительным, однако чем дальше он проникает, тем эффект все больше рассеивается... все больше рассеивается по мере увеличения расстояния. И если вам важна скорость, то вам потребуется именно этот электротонический потенциал. Как только это происходит, его воздействию подвергаются все остальные отделы клетки, однако если вы хотите, чтобы это изменение потенциала распространялось на более длинные расстояния - к примеру, давайте решим, что если мы прошли весь путь до этой точки нейрона и хотим теперь измерить его, то он не будет оказывать какого-либо влияния. Возможно, несколько позднее, однако этот потенциал не будет иметь никакого влияния, поскольку заряд рассеивается по мере того как увеличивается заряд во всей клетке целиком. Итак, это влияние вдали от первоначальной локализации, где произошло открытие ворот. Оно будет существенно меньше. Расстояние на самом деле не идет на пользу действию. И теперь давайте постараемся решить, что будет происходить с потенциалом действия. Из названия ясно, что в этом случае будет больше действия. Итак, давайте начнем с той же ситуации. У нас имеются натриевые ворота, которые открываются под действием определенных стимулов. И сейчас я изображу две мембраны. Вот здесь внешняя сторона. И, соответственно, это - внутренняя сторона. Давайте я изображу - возможно, мы уже сталкивались с этим - но сейчас мы разберем это более подробно. Допустим, это - аксон, и давайте я нарисую здесь еще одни натриевые ворота. Они находятся совсем рядом. И они должны быть трансформирующимися. Итак, они трансформируются, а здесь у меня располагаются еще одни натриевые ворота. Нарисую и их. Но не думаю, что нужно рисовать их в большом количестве. Нужно изобразить одно скопление, чтобы вы понимали, что происходит. Давайте я нарисую еще одни калиевые ворота. Хорошо. Все ворота нарисованы. Давайте договоримся, что они все изначально закрыты. Итак, они все находятся в закрытом положении. И теперь на эти натриевые ворота воздействует стимул. И они открываются. Да, допустим, вот эти ворота открываются. Они стимулируются чем-то и в результате раскрываются. Давайте решим, что конкретно вот эти натриевые ворота открываются под действием определенного стимула, который имеет определенный вольтаж. Пусть ворота открываются когда они достигают значения минус 55 милливольт. Запишу это. И когда они находятся в состоянии покоя, разница потенциалов между внутренним пространством клетки и ее внешним пространством составляет минус 70 милливольт, и они поэтому не открываются. Они остаются закрытыми, однако, если в определенных обстоятельствах этот заряд становится достаточно положительным, чтобы обеспечить значение минус 55 милливольт, эти ворота открываются. Давайте запишем несколько правил, которые определяют то, что происходит с этими воротами. Они закрываются - и все это просто приблизительные числа, главная цель заключается в том, чтобы вы уловили основную идею. Пусть они закрываются при при плюс 35 милливольтах. А наши калиевые ворота открываются при плюс 40 милливольтах, просто чтобы вы уловили основную идею. А закрываются калиевые ворота, при минус 80 милливольтах. Пусть будет так. И что же происходит в результате? Давайте решим, что, по какой-то причине, вольтаж здесь становится минус 55 милливольт. Я сейчас изображу схему, по аналогии с тем, как я это делал раньше. Итак, мне нужно немного пространства, чтобы нарисовать свою схему. Итак, схема та же. Вот это вольтаж мембраны. Хорошо. А вот это время. И давайте представим, что мы измеряем вольтаж - давайте решим, что это мембранный вольтаж вблизи натриевых ворот, расположенных вот здесь. Итак, мы измеряем вольтаж с обеих сторон мембраны. И если отсутствует стимуляция, то мы получаем значение порядка минус 70 милливольт - и вдруг какой-то стимул, по какой-то причине, делает этот участок более положительно заряженным. Возможно, это определенный тип электротонического эффекта, который делает этот участок более положительно заряженным. Возможно, здесь имеются определенные положительные заряды. И, в итоге, этот участок становится более положительно заряженным. Затем натрий-калиевые насосы выкачивают ионы наружу, в результате чего не достигается пороговое значение минус 55 милливольт, в результате ничего не происходит, согласны? Однако когда имеется другой электротонический стимул, или, может быть, несколько, здесь концентрируется большое количество положительных зарядов, в результате чего достигается значение минус 55 милливольт. Запомните, что как только появляется положительный заряд, все становится менее отрицательно заряженным. И разница потенциалов становится меньше. И когда мы достигаем значения минус 55 милливольт - ворота открываются. Вот они были закрыты прежде. Они были закрыты при значении минус 70 милливольт. Итак, давайте я запишу вот здесь. В данной точке, наши натриевые ворота открываются. И теперь что же происходит, когда наши натриевые ворота открываются? Когда они открываются - мы уже наблюдали это раньше - все положительно заряженные ионы натрия направляются вот сюда, как в направлении электрического градиента, так и градиента диффузии, и проникают внутрь клетки. Здесь снаружи имеется такое большое количество натрия, здесь настолько положительный заряд, что они просто стремятся проникнуть внутрь. И как только они достигают этого порога, даже несмотря на то, что это может произойти только при минус 55 или, возможно, при минус 50 милливольтах, в результате ворота открываются и весь наш положительный заряд поступает внутрь клетки. И разница потенциалов становится намного более положительной. Хорошо, теперь дальше. Они продолжают поступать внутрь, и заряд становится все более и более положительным, и по мере того как он становится более положительным, ворота закрываются при значении плюс 35 милливольт. Сейчас освобожу побольше места, чтобы продолжить. Итак, давайте представим, что мы находимся вот здесь - давайте решим, что вот здесь у нас плюс 35 милливольт. Ворота закрываются, и в то же время, все это, что я только что удалил - я установил на значении плюс 40 милливольт, хотя нет, плюс 35, просто чтобы поддержать свою идею. Итак, давайте представим, что при плюс 35 милливольтах наши натриевые ворота открываются. Что же происходит в результате? Внезапно мы оказываемся при плюс 35, или, возможно, при плюс 40 милливольтах вот так - давайте решим, что плюс 40, я полагаю, вы уловили идею так или иначе, поэтому, давайте решим, что плюс 40 . Итак, при плюс 40, эти ворота закрываются. Больше никакие положительно заряженные ионы не поступают внутрь, однако теперь у нас внутри заряд более положительный, по меньшей мере, локально в данной точке на мембране, чем снаружи. И эти ворота открываются. Итак, в результате наши калиевые ворота открываются. K-плюс ионные ворота открываются. И что же происходит теперь, когда они открываются? У нас имеются здесь все эти ионы натрия. Мы уже видели на примере натрий- калиевого насоса, что калий - все эти ионы калия у нас расположены вот здесь. Мы видели на примере натрий-калиевого насоса, что он увеличивает концентрацию натрия на внешней стороне, а концентрация калия при этом выше внутри клетки. И теперь, в дополнении к этому положительному заряду в плюс 40 милливольт, у нас также имеется более положительный заряд на внутренней поверхности мембраны. Итак, они открываются. И эти частицы хотят уйти, поскольку здесь снаружи меньше ионов калия. И они хотят идти по своему градиенту концентрации. На этой стороне также имеется большой положительный заряд. Примерно плюс 40 милливольт. Ионы хотят высвободиться. Они начинают выходить из клетки. Итак, положительные заряды начинают покидать клетку изнутри наружу. И мы снова становимся менее положительно заряженными. Давайте я запишу, что же здесь происходит. Итак, в данной точке наши натриевые ворота закрываются, а открываются калиевые ворота. Одни закрылись, другие открылись. После этого положительные заряды начинают выходить из клетки повторно, и возможно, я несколько сгустил краски, потому что они закроются, вероятно, когда мы уже получим минус 80 милливольт. Допустим, калиевые ворота закроются при -80. И затем наш натрий-калиевый насос может вернуть нас обратно к нашему значению минус 70 милливольт. Итак, вот что происходит на данном участке клетки, в непосредственной близости от первых натриевых ворот. Однако что же происходит, в целом? По мере того как здесь формируется очень пложительный заряд - мы приблизились к значению 40 милливольт вот здесь. Мы достигли 40 милливольт на данном участке клетки. Как вы уже практически проследили это на коротком расстоянии электротонического потенциала, этот участок становится более положительно заряженным. Он становится более положительно заряженным. Эти положительные заряды начинают распространяться туда, где заряд менее положительный. То есть он становится более положительным. Он был при минус 70 милливольт, однако он становится более положительным. Постепенно он увеличивается до минус 65, минус 60, минус 55 и затем «Бам!». Опять происходит стимуляция. И эти ворота снова открываются. Эти ворота опять открываются. Натрий опять поступает внутрь. И если вы хотите проследить за этими воротами, за разницей потенциалов того, что проходит через них, все это происходит как только натрий начинает поступать через эти первые ворота, через вторые ворота - он потенцируется здесь, потому что вторые ворота расположены чуть позже по времени - потому что весь этот поток проходит чуть левее от него, его потенциал увеличивается. Он потенцируется, и с ним происходит снова то же самое. Когда натрий поступает сюда, среда становится положительно заряженной, и это приводит к тому, что клетка, вольтаж вокруг, заряд становятся несколько более положительными, и это запускает открытие следующих натриевых ворот и затем происходит все то же самое, повторяется тот же цикл. Затем калиевые ворота открываются, чтобы отрицательный заряд вернулся, однако к тому времени, как это происходит, среда становится уже достаточно положительно заряженной, чтобы запустить еще одни натриевые ворота. Итак, одни за другими, эти натриевые ворота открываются и закрываются, таким образом передается информация, происходит передача этого изменения потенциала. Итак, что же происходит здесь? Итак, это происходит медленнее и с привлечением энергии. Это происходило - электротонический потенциал - очень быстро. А этот процесс медленный. Потенциал действия более медленный. Я не имею в виду, что он совсем медленный. Он медленнее, потому что он должен задействовать все эти открывания и закрывания ворот, и он также требует энергии. Запишу это. И вы также должны постоянно обеспечивать энергией потенциал в нашей клетке, и использовать ваши натрий-калиевые насосы, которые находятся в активном состоянии. Но это хорошо. Положительной стороной является то, что с его помощью хорошо охватывается расстояние. Это тоже запишем. Мы наблюдали на примере электротонического потенциала, что по мере того, как мы продвигаемся все дальше и дальше от того места, где произошла стимуляция, изменение потенциала становится все более рассеянным. Оно экспоненциально угасает. Оно становится все более рассеянным по мере того, как мы продвигаемся все дальше, что не очень хорошо для больших расстояний. Это же может продолжаться бесконечно, потому что каждый раз когда стимулируются новые ворота, и эти ворота продолжают обеспечивать поток ионов, входящих ионов, а также тех, которые делают так, что среда становится несколько более отрицательно заряженной. Вслед за этим происходит открытие следующих ворот. И это позволяет очень эффективно проходить длинные расстояния. И теперь у нас действительно есть основа для понимания того, что происходит в нейроне, и я продолжу эту тему в следующем видеоролике, где покажу вам, как электротонические потенциалы и потенциалы действия могут объединяться для обеспечения прохождения сигнала по нейрону.
Фазы потенциала действия
- Предспайк - процесс медленной деполяризации мембраны до критического уровня деполяризации (местное возбуждение, локальный ответ).
- Пиковый потенциал, или спайк , состоящий из восходящей части (деполяризация мембраны) и нисходящей части (реполяризация мембраны).
- Отрицательный следовой потенциал - от критического уровня деполяризации до исходного уровня поляризации мембраны (следовая деполяризация).
- Положительный следовой потенциал - увеличение мембранного потенциала и постепенное возвращение его к исходной величине (следовая гиперполяризация).
Общие положения
Поляризация мембраны живой клетки обусловлена отличием ионного состава с её внутренней и наружной стороны. Когда клетка находится в спокойном (невозбуждённом) состоянии, ионы по разные стороны мембраны создают относительно стабильную разность потенциалов, называемую потенциалом покоя . Если ввести внутрь живой клетки электрод и измерить мембранный потенциал покоя, он будет иметь отрицательное значение (порядка −70 - −90 мВ). Это объясняется тем, что суммарный заряд на внутренней стороне мембраны существенно меньше, чем на внешней, хотя с обеих сторон содержатся и катионы , и анионы . Снаружи - на порядок больше ионов натрия , кальция и хлора , внутри - ионов калия и отрицательно заряженных белковых молекул, аминокислот, органических кислот, фосфатов , сульфатов . Надо понимать, что речь идёт именно о заряде поверхности мембраны - в целом среда и внутри, и снаружи клетки заряжена нейтрально.
Потенциал мембраны может изменяться под действием различных стимулов. Искусственным стимулом может служить электрический ток , подаваемый на внешнюю или внутреннюю сторону мембраны через электрод. В естественных условиях стимулом часто служит химический сигнал от соседних клеток, поступающий через синапс или путём диффузной передачи через межклеточную среду. Смещение мембранного потенциала может происходить в отрицательную (гиперполяризация ) или положительную (деполяризация ) сторону.
В нервной ткани потенциал действия, как правило, возникает при деполяризации - если деполяризация мембраны нейрона достигает некоторого порогового уровня или превышает его, клетка возбуждается, и от её тела к аксонам и дендритам распространяется волна электрического сигнала. (В реальных условиях на теле нейрона обычно возникают постсинаптические потенциалы, которые сильно отличаются от потенциала действия по своей природе - например, они не подчиняются принципу «всё или ничего». Эти потенциалы преобразуются в потенциал действия на особом участке мембраны - аксонном холмике, так что потенциал действия не распространяется на дендриты).
Это обусловлено тем, что на мембране клетки находятся ионные каналы - белковые молекулы, образующие в мембране поры, через которые ионы могут проходить с внутренней стороны мембраны на наружную и наоборот. Большинство каналов ионо-специфичны - натриевый канал пропускает практически только ионы натрия и не пропускает другие (это явление называют селективностью). Мембрана клеток возбудимых тканей (нервной и мышечной) содержит большое количество потенциал-зависимых ионных каналов, способных быстро реагировать на смещение мембранного потенциала. Деполяризация мембраны в первую очередь вызывает открытие потенциал-зависимых натриевых каналов. Когда одновременно открывается достаточно много натриевых каналов, положительно заряженные ионы натрия устремляются через них на внутреннюю сторону мембраны. Движущая сила в данном случае обеспечивается градиентом концентрации (с внешней стороны мембраны находится намного больше положительно заряженных ионов натрия, чем внутри клетки) и отрицательным зарядом внутренней стороны мембраны (см. Рис. 2). Поток ионов натрия вызывает ещё бо́льшее и очень быстрое изменение мембранного потенциала, которое и называют потенциалом действия (в специальной литературе обозначается ПД).
Согласно закону «всё-или-ничего» мембрана клетки возбудимой ткани либо не отвечает на стимул совсем, либо отвечает с максимально возможной для неё на данный момент силой. То есть, если стимул слишком слаб и порог не достигнут, потенциал действия не возникает совсем; в то же время, пороговый стимул вызовет потенциал действия такой же амплитуды , как и стимул, превышающий пороговый. Это отнюдь не означает, что амплитуда потенциала действия всегда одинакова - один и тот же участок мембраны, находясь в разных состояниях, может генерировать потенциалы действия разной амплитуды.
После возбуждения нейрон на некоторое время оказывается в состоянии абсолютной рефрактерности , когда никакие сигналы не могут его возбудить снова, затем входит в фазу относительной рефрактерности , когда его могут возбудить исключительно сильные сигналы (при этом амплитуда ПД будет ниже, чем обычно). Рефрактерный период возникает из-за инактивации быстрого натриевого тока, то есть инактивации натриевых каналов (см. ниже).
Распространение потенциала действия
По немиелинизированным волокнам
По ходу ПД каналы переходят из состояния в состояние: у Na + каналов основных состояний три - закрытое, открытое и инактивированное (в реальности дело сложнее, но этих трёх достаточно для описания), у K + каналов два - закрытое и открытое.
Поведение каналов, участвующих в формировании ПД, описывается через проводимость и высчиляется через коэффициенты переноса (трансфера).
Коэффициенты переноса были выведены Ходжкином и Хаксли.
Проводимость для калия G K на единицу площади
| d n / d t = α n (1 − n) − β n n {\displaystyle dn/dt=\alpha _{n}(1-n)-\beta _{n}n} , |
| где: |
| α n {\displaystyle \alpha _{n}} - коэффициент трансфера из закрытого в открытое состояние для K+ каналов ; |
| β n {\displaystyle \beta _{n}} - коэффициент трансфера из открытого в закрытое состояние для K+ каналов ; |
| n {\displaystyle n} - фракция К+ каналов в открытом состоянии; |
| (1 − n) {\displaystyle (1-n)} - фракция К+ каналов в закрытом состоянии |
рассчитывается сложнее, поскольку, как уже было сказано, у потенциал-зависимых Na+ каналов, помимо закрытого/открытого состояний, переход между которыми описывается параметром m {\displaystyle m} , есть ещё инактивированное/не-инактивированное состояния, переход между которыми описывается через параметр h {\displaystyle h}
| d m / d t = α m (1 − m) − β m m {\displaystyle dm/dt=\alpha _{m}(1-m)-\beta _{m}m} , | d h / d t = α h (1 − h) − β h h {\displaystyle dh/dt=\alpha _{h}(1-h)-\beta _{h}h} , |
| где: | где: |
| α m {\displaystyle \alpha _{m}} - коэффициент трансфера из закрытого в открытое состояние для Na+ каналов ; | α h {\displaystyle \alpha _{h}} - коэффициент трансфера из инактивированного в не-инактивированное состояние для Na+ каналов ; |
| β m {\displaystyle \beta _{m}} - коэффициент трансфера из открытого в закрытое состояние для Na+ каналов ; | β h {\displaystyle \beta _{h}} - коэффициент трансфера из не-инактивированного в инактивированное состояние для Na+ каналов ; |
| m {\displaystyle m} - фракция Na+ каналов в открытом состоянии; | h {\displaystyle h} - фракция Na+ каналов в не-инактивированном состоянии; |
| (1 − m) {\displaystyle (1-m)} - фракция Na+ каналов в закрытом состоянии | (1 − h) {\displaystyle (1-h)} - фракция Na+ каналов в инактивированном состоянии. |
Соединение в живых существах органической материи и металлов, использование живой клетки в качестве проводника электрического тока не могут не удивлять. Чем больше подробностей о своем теле я узнаю, тем чаще возникает удивление от простых его действий. Сгибание руки может восприниматься не как привычный, почти не замечаемый процесс, а как результат слаженной работы мышц и сухожилий, получивших команду от мозга, переданную по нервным волокнам с помощью электричества. Разглядывая полосу глубоко переливающейся синевы, возникающей на куске полированного лабрадорита, который я верчу в руках, я воспринимаю излучаемые им электрические волны рецепторами глаз. В рецепторах эта информация кодируется в виде череды электрических импульсов. Она передается в головной мозг, обрабатывается им, а результат всего этого — восприятие таинственно-красивого камня, возникающие при этом эмоции. Матрица, короче:)
Вместе с восхищением возникает и желание узнать — как это все происходит, желание вникать все глубже в подробности каждого процесса.
Потенциал действия — это тот электрический импульс, которой доставляет информацию от рецепторов к мозгу, и с помощью которого мозг управляет телом. Для его существования необходимы особо устроенные клетки, нейроны, мембраны которых облеплены с обеих сторон положительно и отрицательно заряженными ионами, которые только и ждут сигнала, чтобы этот электрический импульс пронести по всей длине клетки. Основным местом существования потенциала действия является аксон нейрона. Дендриты некоторых типов нейронов также способны проводить электрический импульс.

Достаточно взглянуть на картинку слева, чтоб увидеть основное отличие потенциала покоя от потенциала действия: когда мембрана находится в состоянии покоя, на всей ее протяженности снаружи существует положительный электрический заряд, а внутри — отрицательный.

А потенциал действия, проходящий по нейрону от тела клетки до конца аксона, представляет собой изменение внешнего и внутреннего зарядов клеточной мембраны на небольшом участке нейрона: на очень краткий промежуток времени отрицательный и положительный заряды по разным сторонам мембраны меняются местами.
Возникают вопросы: что запускает потенциал действия? Какова его физическая природа, что именно происходит в клетке при его существовании? Каким образом он распространяется? Почему он идет только в одном направлении? Как клетка восстанавливает исходный потенциал покоя после прохождения по ней волны возбуждения?
Итак, первый вопрос: как возникает потенциал действия? Так как передача сигналов от нейронов-источников к нейрону-получателю, или постсинаптическому нейрону (postsynaptic neuron ), происходит в синапсах, запускающий потенциал действия сигнал стоит искать именно там. Через синапс в нейрон приходят сигнальные вещества, но их поступление не обязательно означает запуск возбуждения нейрона. Если бы нейрон реагировал на очень малые количества сигнальных веществ, это было бы так же неудобно, как иметь супер-чувствительный выключатель, срабатывающий, если на него случайно чихнуть.

Необходимо достаточно сильное воздействие определенных сигнальных веществ, чтобы запустить потенциал действия. В реальных условиях на теле нейрона обычно возникают постсинаптические потенциалы — временные изменения зарядов мембраны, вызываемые пришедшими через синапс сигнальными веществами .
Постсинаптические потенциалы сильно отличаются от потенциала действия по своей природе. Если потенциал действия имеет одну строго определенную величину, то величина постсинаптических потенциалов зависит только от вида и количества вызвавших их химических веществ, переданных через синапсы. В то время как потенциалы действия, генерируемые разными нейронами, примерно одинаковы, постсинаптические потенциалы, возникающие в разных входных синапсах на одном и том же нейроне, сильно отличаются по величине и продолжительности. В одном синапсе нейрона приходящий нервный импульс может вызвать изменение мембранного потенциала на 0,1 мВ, а в другом — на 20 мВ. Кроме того, существуют как вызывающие потенциал действия, так и тормозящие его сигналы.
Тело клетки служит своеобразным калькулятором, простейшим аналоговым компьютером, суммирующим все поступающие в него сигналы в виде возникающих постсинаптических потенциалов. Аналоговые компьютеры устроены очень просто. Например можно построить водяной аналоговый компьютер. Если из одной трубы в банку вливается три килограмма воды, а из другой в ту же банку — пять, то взвесив банку и выяснив, что весы показывают вес, равный восьми килограммам, мы делаем вывод, что 3+5=8. На заре развития компьютерной техники использовались и водяные, и прочие аналоговые вычислительные машины, но наиболее продвинутой была, конечно, машина, которая в точности как и нейроны суммирует электрические токи. Операторы таких ЭВМ занимались вычислениями, перетыкая штекеры из одного гнезда в другое, сливая вместе разные токи.
Так вот в нейроне происходит такое же суммирование электрических токов. В процессе этого суммирования может быть перейден порог возбуждения (excitation threshold ): будет достигнуто определенное значение мембранного потенциала, которое запускает потенциал действия. Обрати внимание на то, что для достижения порога возбуждения мембранной потенциал должен увеличиваться. Если мембранный потенциал покоя нейрона равен -70 мВ, то его порог возбуждения может быть равен -55 мВ. Снижение мембранного потенциала например до -90 мВ приводит к тому, что для преодоления порога возбуждения требуется более сильное воздействие, ведь он остается прежним: -55мВ.

В постсинаптическом нейроне, получающем сигналы от двух возбуждающих и одного тормозящего нейронов, могут существовать различные комбинации сигналов, и далеко не каждая из них приводит к возникновению потенциала действия.
В первой схеме картинки вверху изображена ситуация, когда только от одного возбуждающего нейрона приходит слабый сигнал, которого недостаточно для преодоления порога возбуждения. Этот сигнал повторяется, но к моменту прихода второго, такого же слабого сигнала от того же аксона возбуждение мембраны уже достигает потенциала покоя, и потенциал действия не возникает.
На второй схеме изображен тот же самый слабый сигнал, но приходящий с большей частотой. Именно из-за увеличения частоты сигнала мембрана не успевает вернуться в состояние покоя в промежутке между двумя сигналами. В этом случае сумма остаточного заряда мембраны после первого сигнала и поступающего следом второго сигнала позволяет преодолеть порог возбуждения нейрона. В этом случае у основания аксона, которое называется аксонным холмиком (axon hillock [‘aksɒn ‘hɪlək] ), возникает потенциал действия.
На третьей схеме от обоих возбуждающих нейронов приходят слабые сигналы, и их суммы достаточно для запуска потенциала действия.
Четвертая схема показывает воздействие на постсинаптический нейрон возбуждающего и тормозящего сигнала. При поступлении только возбуждающего сигнала мембранный потенциал незначительно увеличивается, а при поступлении только тормозного сигнала — снижается. Если эти два сигнала приходят одновременно, то они всего лишь уничтожают друг друга, оставляя мембранный потенциал в состоянии, близком к потенциалу покоя. Это напоминает надавливание на одну и ту же дверь с разных сторон: по отдельности каждое воздействие могло бы приоткрыть дверь или наоборот, прикрыть ее, но существующие одновременно, они приводят лишь к небольшому сдвигу ее по направлению действия большей по модулю силы.
Природа системы такова, что даже малые по величине постсинаптические потенциалы, суммируясь, могут давать большой эффект. Нейрон «складывает» все поступившие в него возбуждающие постсинаптические потенциалы, «вычитает» поступившие тормозные постсинаптические потенциалы, и как только будет перейден порог возбуждения, возникает потенциал действия.
Чтобы разобраться, что происходит в клетке во время запуска и прохождения потенциала действия , необходимо перечислить каждого участвующего игрока и его действие, как было сделано при описании возникновения мембранного потенциала покоя в . Итак, действующие лица при создании потенциала действия :

1 . , которые были закрыты при существовании мембранного потенциала покоя, открываются, как только потенциал достигает порога возбуждения. Так как внутри клетки на мембране существует отрицательный заряд, то в этот момент через каналы туда врываются притягиваемые им
2 . положительно заряженные ионы натрия (Na +). Именно они создавали положительный заряд на внешней поверхности мембраны нейрона, и именно в этот момент они переносят его внутрь, оставляя на внешней мембране отрицательный заряд, который создают находящиеся вне клетки отрицательно заряженные ионы хлора.
3 . таким образом ионы хлора (Cl —) создают отрицательный заряд на внешней поверхности мембраны.

Перемещение ионов натрия внутрь клетки приводит к резкому изменению зарядов внутри и снаружи нейрона. Место существующего во время покоя нейрона отрицательного внутреннего заряда мембраны занимает положительный заряд ионов натрия. А снаружи клеточной мембраны после ухода оттуда Na + наконец-то может проявить себя Cl — , чей отрицательный заряд больше не компенсируется положительно заряженными ионами. Таким образом потенциал клеточной мембраны внезапно возрастает с -70 до +40 мВ. Именно это повышение, называемое деполяризацией (depolarization ), изображено резко идущей вверх частью пика на этой картинке. Такое повышение мембранного потенциала является сигналом закрыть потенциал-зависимые натриевые каналы, а также открывает потенциал-зависимые калиевые каналы, о которых раньше мы еще не говорили.
4 . потенциал-зависимые калиевые каналы. Они существуют в нейроне в дополнение к проточным калиевым каналам, которые, как ты наверное помнишь из прошлой главы про потенциал, не имеют ворот и обеспечивают постоянный отток калия из клетки, играя существенную роль в формировании потенциала покоя. Так вот оказывается, что есть две разновидности калиевых каналов, и сейчас мы рассмотрим работу потенциал-зависимого калиевого канала. Открываясь после того, как потенциал действия достиг своего пика, потенциал-зависимые калиевые каналы резко увеличивают отток ионов калия из клетки. Существующий в этот момент внутри нейрона положительный заряд, образовавшийся за счет притока Na + , активно выпихивает положительно заряженные ионы калия (К +) из клетки через оба вида каналов.

Итак, еще раз проследим цепочку событий до этого момента: потенциал достигает порога возбуждения, и этот электрический сигнал приводит к тому, что открылись потенциал-зависимые натриевые каналы, ионы Na + хлынули внутрь, притягиваемые отрицательно заряженными протеинами, произошла деполяризация, то есть разница зарядов снаружи и внутри резко снизилась, и даже более того — отсутствие положительно заряженных ионов натрия привело к возникновению отрицательного заряда там, где только что был положительный — произошла смена полюсов. Действие электрического импульса закончилось, натриевые каналы вновь закрылись, и столпившиеся внутри ионы Na + вытолкнули одноименно заряженные ионы К + через открывшиеся потенциал-зависимые калиевые каналы и через проточные калиевые каналы.
Заряд внутренней поверхности клеточной мембраны начинает стремительно снижаться, на схеме этому соответствует идущая вниз часть пика. Такое снижение заряда называется реполяризацией (repolarization ), то есть разница зарядов вне и внутри нейрона вновь восстановилась, как и было изначально в состоянии покоя. Но восстановилась именно разница зарядов, а не ионный состав, ее создающий. Несмотря на то, что в конце реполяризации снаружи мембраны образуется положительный заряд, равный положительному заряду мембраны в состоянии покоя, до достижения мембраной состояния покоя еще далеко. Ведь этот положительный заряд принесли туда вышедшие из клетки ионы К + , а при потенциале покоя его поддерживают ионы Na + , которые в данный момент заперты внутри клетки, в том самом месте, где ранее в состоянии покоя находились ионы калия.
В конце реполяризации может возникнуть снижение мембранного потенциала даже ниже его значения для покоящегося нейрона, или гиперполяризация (hyperpolarization [ˌhaɪpəpələraɪ’zeɪʃn] ). Через короткий промежуток времени она компенсируется дополнительным выходом ионов калия из клетки, и наступает прежнее, стабильное состояние покоя.

Когда мембранный потенциал становится равен потенциалу покоя (-70 мВ), потенциал-зависимые калиевые каналы закрываются. Через проточные калиевые каналы К + уже не вытекает так стремительно, ведь его концентрация внутри клетки упала, а снаружи находится много ионов калия. В то же время внутри находится много ионов натрия, которые должны быть выведены из клетки для того, чтоб через нее мог снова пройти потенциал действия.
Восстановлением исходного положения «натрий снаружи, калий внутри» занимается натрий-калиевый насос, выводящий из клетки три Na + и одновременно проносящий внутрь два К + . О его работе было рассказано в предыдущей главе про потенциал, и он схематично показан в четвертой части этой картинки. Совместные действия натрий-калиевого насоса и проточных калиевых каналов приводят к тому, что с внешней стороны мембраны опять возникает большая концентрация Na + , а концентрация К + внутри нейрона выше наружной.
Если бы работали только проточные калиевые каналы, то ионы калия только выходили бы из клетки, и было бы невозможным поддерживать внутри большую концентрацию К + , чем снаружи. Если бы действовали только потенциал-зависимые натриевые каналы, то ионы натрия вообще не могли бы покинуть клетку после того, как вошли туда, создавая потенциал действия. Но натрий-калиевый насос (ну то есть натрий-калиевый канал) решает обе эти проблемы: он выводит наружу Na + , восстанавливая его высокую концентрацию снаружи. И загоняет внутрь прошедшие через проточные каналы К + , не давая им возможности распределиться с обеих сторон мембраны равномерно, лишившись таким образом парциального давления, необходимого для формирования мембранного потенциала покоя.
Через тысячную долю секунды после того, как реполяризация нейронной мембраны достигла минимума, активная работа мембранных каналов восстанавливает потенциал покоя. Нейрон готов к проведению следующего потенциала действия.
5 . положительно заряженные ионы кальция (Ca 2+) играют важную роль в формировании потенциала действия лишь в некоторых нервных клетках, поэтому в данной главе, содержащей общую для различных нейронов информацию, она не будет описана.

На этом общее описание перемещения ионов через клеточную мембрану, превращающее потенциал покоя нейрона в потенциал действия, закончено. Можно сделать перерыв и переварить его вместе с чем-нибудь вкусным, заодно подпитав мозг, компенсировав те калории, которые он затратил на работу по обработке всей этой информации.
Распространение потенциала действия по всей длине аксона обеспечивается тем фактом, что возникший при его существовании в одном участке мембраны нейрона электрический ток возбуждает соседние участки, открывая натриевые потенциал-зависимые каналы и запуская уже там потенциал действия. Точнее, электрический ток запускает потенциал действия только с одной стороны от места существования предыдущего потенциала. Потому что с другой стороны натриевые каналы инактивированы после недавнего открытия, а без их участия возникновение потенциала действия невозможно. Если бы не было этого временного промежутка нефункциональности между двумя открытиями натриевых каналов, потенциал действия не мог бы распространяться только в одном направлении. Происходящая в результате цепная реакция открытия натриевых каналов обеспечивает передачу нервного импульса от места его возникновения до окончаний аксона, подходящих к другим нейронам (или к мышечной клетке).

Схематично эту цепную реакцию можно изобразить так:
Момент времени 1 : Электрический ток, возникший на в результате открытия потенциал-зависимых натриевых каналов на участке 1 , возбуждает соседние участки мембраны.
Момент времени 2 : В результате этого возбуждения на участке 2 открываются потенциал-зависимые натриевые каналы и возникает потенциал действия, который, в свою очередь, возбуждает соседние участки мембраны. На участке 1 открылись потенциал-зависимые калиевые каналы, восстанавливающие мембранный потенциал после прохождения потенциала действия.
Момент времени 3 : Электрический ток, возникший в результате открытия потенциал-зависимых натриевых каналов на участке 2 , вызвал открытие таких же каналов на участке 3 , и потенциал действия перешел туда. На участке 1 потенциал-зависимые натриевые каналы не смогли открыться, несмотря на электрическое возбуждение от участка 2 , так как они были временно инактивированы после создания потенциала действия. На участке 2 открылись потенциал-зависимые калиевые каналы, восстанавливающие мембранный потенциал покоя после прохождения потенциала действия.
Мы уже знаем, что отростки нейронов могут быть покрыты обернутыми вокруг них клетками нейроглии, или миелиновой оболочкой. Именно она делает аксон похожим на вытянутую цепочку сосисок, про ее образование и функции говорилось в . Передача потенциала возбуждения по «голым», не покрытым миелиновыми клетками, нервным волокнам отличается от его прохождения по миелинизированному аксону. При сравнении заметны существенные преимущества, предоставляемые обернутыми вокруг аксона клетками нейроглии:

1 . Экономичность . Мембрана аксона под миелином не имеет каналов для пропускания ионов, создающих потенциал действия. Каналы существуют только в узких промежутках между глиальными клетками, которые называют перехваты Ранвье (Ranvier’s constriction ), и только на этих узких участках аксона возникает потенциал действия. Концентрация ионных каналов в этих перехватах в 100 раз выше, чем в мембранах безмиелиновых волокон, тем не менее для того, чтобы обеспечить их работу, требуется значительно меньше энергии по сравнению с тем, сколько энергии потребовалось бы для обеспечения ионных каналов, расположенных по всей поверхности «голого» аксона. Кроме того, такая экономичность позволяет быстрее «перезаряжать» аксон для проведения следующего потенциала действия, поэтому миелинизированные волокна способны передавать сигналы более высокой частоты.
2
. Скорость
. По «голому» аксону сигнал передается в каждой его точке. Ионные каналы открываются в каждой точке его поверхности, один за другим, что напоминает тщательное, скрупулезное вчитывание в текст.
В миелинезированном аксоне потенциал действия, возникший в одном перехвате Ранвье, своим электрическим полем дотягивается до соседних перехватов, что приводит к возникновению в них новых потенциалов действия. То есть возбуждение переходит скачкообразно, от одного перехвата к другому. В случае повреждения одного перехвата Ранвье потенциал действия возбуждает следующий после поврежденного перехват, а в случае необходимости даже третий, четвертый и пятый по счету. Распространение электрического поля на такую длину возможно из-за электроизоляции, создаваемой миелиновыми муфтами: она уменьшает рассеивание электрического поля, которое неизбежно для безмиелиновых волокон.
Таким образом, скорость распространения потенциала действия по миелинизированным волокнам намного выше по сравнению с немиелинизированными. Ведь за один и тот же отрезок времени потенциал действия на «голом» аксоне возбудит лишь находящийся непосредственно рядом участок, а потенциал действия на миелинизированном волокне перепрыгнет на расстояние, равное расстоянию между одним или несколькими перехватами Ранвье:


Диапазон скоростей передачи потенциала действия в различных миелинизированных волокнах очень широк: от нескольких метров в секунду до «мирового рекорда», установленного аксоном креветки: он проводит возбуждение со скоростью, превышающей 200 м/с.
Чтобы представить, насколько эффективно может быть увеличена скорость проведения потенциала действия за счёт миелиновой оболочки, достаточно сравнить скорость его распространения по немиелинизированным и миелинизированным волокнам нейронов человека. Если взять скорость проведения импульса в одинаково тонких волокнах, то в безмиелиновом волокне она составит примерно 1 м/с, а в миелинизированном 15-20 м/с. В толстых аксонах человека, покрытых миелиновой оболочкой, скорость проведения может достигать 120 м/с.
Итак, подведем итог : передача электрического импульса через тело и отростки нейрона осуществляется с помощью прохождения ионов через клеточную мембрану.

1 . сначала на клеточной мембране создается потенциал покоя . Его образуют положительно заряженные ионы натрия (Na + ) снаружи клетки и отрицательно заряженные протеины внутри клетки. Создающие заряды частицы расположены очень близко к клеточной мембране, ведь они имеют разные знаки и притягиваются друг к другу. Поэтому потенциал покоя существует только возле мембраны, в целом среда внутри нейрона и вне его электрически нейтральна. Кроме Na + и отрицательно заряженных протеинов, в создании и поддержании электрического заряда участвуют положительно заряженные ионы (К + ) и отрицательно заряженные ионы хлора (Cl — ) , тоже находящиеся вблизи мембраны. Потенциал покоя равен примерно -70 мВ (в дальнейшем при указывании численного значения мембранных потенциалов слово «примерно» будет опускаться).
2 . когда нейрон через синапсы получает сигналы от других нейронов, под их влиянием на мембране тела нейрона начинают возникать постсинаптические потенциалы , то есть изменения величины электрических зарядов на внутренней и внешней поверхностях мембраны. Под влиянием этих сигналов внутренний отрицательный заряд и внешний положительный могут увеличиваться или уменьшаться на разные величины. И нейрон выступает в роли калькулятора, суммирующего эти воздействия.

3 . итогом суммирования постсинаптических потенциалов может являться продолжение нахождения нейрона в состоянии покоя. Но если сумма дойдет до значения, называемого порогом возбуждения , то в аксонном холмике (утолщении тела нейрона у основания аксона) возникнет потенциал действия .
4 . в создании потенциала действия главную роль играет клеточная мембрана . Она регулирует прохождение через нее ионов (всех перечисленных, кроме отрицательно заряженных протеинов, которые не могут покидать пределы клетки), открывая и закрывая специализированные каналы, способные проводить только определенные ионы.
5 . когда мембрана нейрона переходит порог возбуждения, в ней открываются потенциал-зависимые натриевые каналы . Исчезает то препятствие, которое отделяло положительно заряженные Na + от притягивающих их отрицательно заряженных протеинов, и Na + устремляются внутрь клетки . Таким образом наружный положительный заряд переносится внутрь клетки, а снаружи остается отрицательный заряд, создаваемый ионами Cl — . Происходит изменение мембранного потенциала от потенциала покоя, равного -70 мВ, до +40 мВ, и это называют потенциалом действия . Важно понимать, что это изменение потенциала происходит только на том участке мембраны, где открылись потенциал-зависимые натриевые каналы.
6 . В момент достижения мембраной потенциала в 40 мВ потенциал-зависимые натриевые каналы закрываются, и на этом же участке открываются потенциал-зависимые калиевые каналы . Снаружи в этот момент существует отрицательный заряд, а внутри — положительный, создаваемый вошедшими туда ионами Na + и уже находящимися там в большом количестве ионами К + . Ионы калия выпихиваются в окружающую нейрон среду через открывшиеся каналы, что резко снижает внутренний заряд мембраны и увеличивает внешний. По обеим сторонам мембраны создаются заряды той же величины, которые обычно поддерживают потенциал покоя. Но это состояние клетки отличается от потенциала покоя тем, что ионы калия и натрия находятся не на своих местах: Na + находится внутри, а К + снаружи. Опять же все описанное касается только одного места на мембране, в котором только что перестал существовать потенциал действия.
7 . Чтобы переместить положительно заряженные ионы Na + и К + по нужные для поддержания потенциала покоя стороны мембраны, используется натрий-калиевый насос . И это действие тоже касается ограниченного участка мембраны: того, где только что закрылись потенциал-зависимые калиевые каналы.
Один цикл работы натрий-калиевого насоса заключается в том, что он захватывает изнутри клетки три иона Na + , перемещает их наружу, где захватывает два иона К + и перемещает их внутрь . То есть канал обменивает три иона Na + на два иона К + , причем Na + могут перемещаться только наружу, а К + только внутрь клетки. Таким образом восстанавливается исходная концентрация натрия на наружной стороне мембраны и калия на внутренней. Натрий-калиевый насос выводит из клетки три положительно заряженных иона, а вводит в нее только два. Следовательно, при каждом его цикле внутренний положительный заряд мембраны уменьшается, а внешний увеличивается. К тому же и вытекающие ионы калия уносят изнутри наружу свои положительные заряды. Возникает вопрос: если потенциал мембраны уже и так был равен потенциалу покоя после входа ионов калия внутрь клетки, зачем его снижать еще сильнее? Ответ заключается в том, что кроме упомянутых здесь ионов, существуют и другие, чье участие в формировании потенциала покоя менее существенно, и именно для компенсации их влияния необходимо такое выравнивание мембранного потенциала. Итогом этого процесса является достижение мембранного потенциала покоя: заряд внутренней стороны мембраны отрицателен, внешней положителен, разность потенциалов равна -70 мВ.
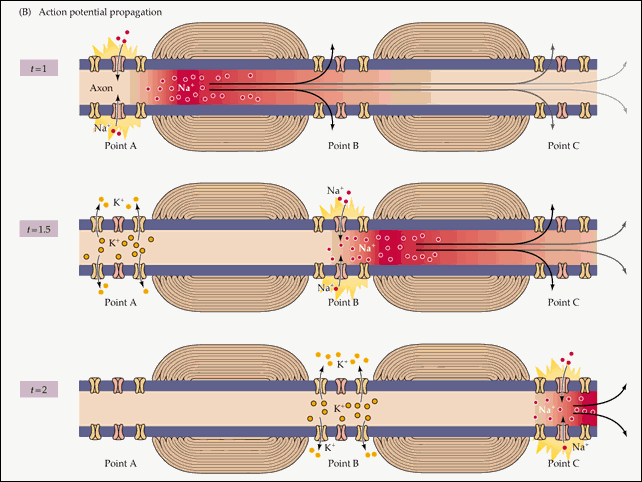
8 . Итак, в том месте клеточной мембраны, где всего три шага назад существовал потенциал действия, опять царит потенциал покоя. Но потенциал действия не прошел бесследно: его существование (описанное в пункте 5) создало электрическое поле, возбудившее соседние с ним части мембраны . С одной стороны (по направлению к концу аксона) это возбуждение вызвало открытие потенциал-зависимых натриевых каналов, запустив уже на новом участке потенциал действия. Который в свою очередь порождает электрическое поле, запускающее следующий потенциал действия, и этот цикл повторяется по всей длине аксона, от его основания до концов, просоединенных к следующим в нервной цепи клеткам.
С другой стороны от потенциала действия, ближе к телу нейрона, несмотря на воздействие электрического поля потенциал действия не возникает. Потому что потенциал-зависимые натриевые каналы на этом участке совсем недавно открывались, и на некоторое время они инактивированы. Именно эта пауза в их работе обеспечивает распространение потенциала действия только в одном направлении.
9 . Любой сигнал, будь то передача информации от сенсорного нейрона в мозг, или команда мозга, идущая к мотонейрону, передается одинаковыми по величине потенциалами действия . Результат потенциала действия (появление восприятия запаха, обусловленное работой сенсорных нейронов, или перемещение тела поближе к его источнику в результате слаженной работы мотонейронов) зависит исключительно от того, в какую нервную цепь включен нейрон, этот потенциал передающий. Для передачи интенсивности сигнала (силы запаха или величины сокращения мышцы) используется частота, с которой потенциалы действия передаются по нейрону. Слабый сигнал передается редкими, а сильный — частыми потенциалами действия .
10 . миелиновая оболочка вокруг отростков нейрона повышает скорость распространения потенциала действия за счет того, что сигнал передается на большое расстояние, равное ширине одной обернутой вокруг отростка клетки нейроглии, а не на непосредственно прилегающий к возбужденному участок, как это происходит у безмиелиновых волокон.
Биологическая реакция
– ответная реакция клеток, тканей, органов в ответ на раздражитель (стимул).
Раздражимость
– свойство всех живых тканей изменять своё внутреннее состояние при изменении внешних условий.
Виды тканей в зависимости от реагирования на внешние раздражители:
I Возбудимые
- обладают свойством возбудимости, т.е. способностью возбуждаться нервная, мышечная, железистая.
II Невозбудимые
– изменяют свое состояние, но не генерируют процесс возбуждения в ответ на нанесенный стимул.
Возбудимость
– способность ткани переходить в возбуждённое состояние.
Возбуждение
– деятельное состояние тканей в ответ на действие раздражителя, это сложная биологическая реакция, проявляющаяся в совокупности физических, физико-химических и функциональных изменений, способная к распространению по ткани.
Возбуждение включает в себя неспецифические и специфические компоненты.
Неспецифические:
сдвиг химических реакций, образование тепла, физико-химические изменения,
продукция биопотенциалов, структурные изменения в мембране клеток.
Специфические:
мышечная ткань отвечает мышечным сокращением, нервная ткань - генерацией нервного импульса и его проведением, железистая ткань – образованием и выделением секрета.
Возбуждение может быть локальным и динамическим (распространяющимся).
Биопотенциалы
Луиджи Гальвани 1791 г. в эксперименте показал, что живые ткани содержат «животное электричество», его научный оппонент, физик Вольта - что это электричество от разнородных металлов, он создал первый источник постоянного тока, который носит название гальванический элемент.
Виды биопотенциалов:
1. Биопотенциал покоя (мембранный) - МПП.
2. Биопотенциал действия (возбуждения) - ПД.
- Биопотенциал покоя – это разность потенциалов между наружной и внутренней поверхностью мембраны клетки в покое. Наружная поверхность мембраны клетки имеет положительный заряд, а внутренняя – отрицательный.
Биопотенциал покоя регистрируется внутриклеточным методом – с помощью микроэлектродов, один из которых вводится внутрь клетки (рис.1).
Рисунок 1. Схематическое представление метода регистрации биопотенциалов.
В эксперименте биопотенциал покоя можно зарегистрировать между повреждённым и неповреждённым участком ткани. Повреждённый участок является моделью внутренней поверхности мембраны клетки.
При внутриклеточном отведении перезарядка мембраны регистрируется под одним электродом (однофазный ПД), при внеклеточном отведении потенциал действия проходит через два электрода (регистрируется двухфазный ПД).
- Биопотенциал действия – это кратковременные высокоамплитудные изменения МПП, которые возникают при возбуждении. ПД регистрируется в раздражаемых тканях, в которых возникает волна возбуждения (рис.2). Измеряется ПД с помощью внутриклеточного отведения и внеклеточного отведения.

Рисунок 2. Потенциал действия, основные его фазы.
Современная, экспериментально доказанная, мембранно-ионная теория возникновения биопотенциалов (Ходжкин, Хаксли, Катц).
Основные положения:
- Электрические процессы возникают на плазматической мембране клетки, которая состоит из бимолекулярного слоя липидов (остов мембраны) и белков, которые выполняют различные функции в мембране: рецепторную, ферментативную, образуют в ней каналы и насосы (рис.3).
Канал мембраны может быть неспецифическим, он постоянно открыт, не имеет воротного механизма, электрические воздействия не изменяют его состояния. Называют каналом «утечки». Специфические каналы (селективные) имеют воротный механизм, поэтому могут находиться или в открытом, или в закрытом состоянии в зависимости от электрических воздействий на мембрану и пропускают только определенный ион. Этот канал состоит из трех частей: водной поры – выстлана внутри гидрофильными группами; селективного фильтра – на наружной поверхности, который пропускает ионы в зависимости от их размера и формы; ворот – на внутренней поверхности мембраны, управляют проницаемостью канала.

Рисунок 3. Строение биологической мембраны.
Канал мембраны может быть неспецифическим, он постоянно открыт, не имеет воротного механизма, электрические воздействия не изменяют его состояния. Называют каналом «утечки». Специфические каналы (селективные) имеют воротный механизм, поэтому могут находиться или в открытом, или в закрытом состоянии в зависимости от электрических воздействий на мембрану и пропускают только определенный ион. Этот канал состоит из трех частей: водной поры – выстлана внутри гидрофильными группами; селективного фильтра – на наружной поверхности, который пропускает ионы в зависимости от их размера и формы; ворот – на внутренней поверхности мембраны, управляют проницаемостью канала (рис.4).

Рисунок 4. Строение ионного канала.
Каналы для натрия имеют два типа ворот: быстрые активационные и медленные инактивационные. В покое открыты медленные инактивационные и закрыты быстрые активационные. При возбуждении происходит открытие быстрых активационных и медленное закрытие медленных инактивационных, т.е. на короткий промежуток времени оба типа ворот открыты (рис.5).

Рисунок 5. Работа активационные и инактивационный воротных механизмов натриевого ионного канала.
Калиевые каналы
имеют только медленные ворота.
Насосы выполняют функцию транспорта через мембрану ионов против градиента концентрации, для их работы используется энергия АТФ.
- По обе стороны мембраны существует концентрационный градиент.
Внутри клетки в 40 раз > К+; t;/p>
Вне клетки: в 20-30 раз > Na+,
в 50 раз > Cl-.
- Мембрана пропускает молекулы жирорастворимых веществ, а анионы органических кислот не проходят. Мембрана проницаема для воды, для ионов проницаемость мембраны различна: для калия в состоянии покоя проницаемость почти в 25 раз больше, чем для натрия. При возбуждении увеличивается проницаемость и для калия (постепенно), и для натрия (быстро, но на очень короткий промежуток времени).
Потенциал покоя
Проницаемость мембраны для ионов К+ повышена, поэтому калий играет основную роль в генерации МПП. Калий создаёт электрическое поле и заряжает наружную поверхность мембраны «+». В тот момент, когда «+» потенциал наружной стороны достигает определённой величины по отношению к «–» внутри, который создается анионами – наступает динамическое равновесие между входящими и выходящими из клетки ионами К+. Этому моменту соответствует потенциал равновесия для К - потенциал покоя.
МПП характеризуется:
1. постоянством;
2. полярностью, снаружи «+», внутри «-»;
3. величина – в мВ, для скелетной мышцы - 60 – 90 мВ,
для гладкой - -30 – 70 мВ,
для нерва -50 – 80мВ,
для секреторной клетки - -20мВ.
МПП - один из основных показателей состояния физиологического покоя клетки. При увеличении внеклеточной концентрации калия уменьшается МПП, т.к. уменьшается диффузия калия из клетки в связи со снижением его концентрационного градиента. При действии веществ, блокирующих ресинтез АТФ, т.к. прекращается работа натрий-калиевого насоса, также снижается МПП. Ионы натрия и хлора входят в клетку, но ввиду низкой проницаемости значительного влияния на МП не оказывают.
Потенциал действия
При возбуждении – резко увеличивается (в несколько тысяч раз) проницаемость для ионов Na, которые поступают внутрь клетки лавинообразно и заряжают внутреннюю сторону «+» - происходит деполяризация мембраны, а затем количество ионов натрия внутри превышает калиевый заряд на поверхности и это приводит к перезарядке мембраны (реверсии). Постепенно увеличивающаяся проницаемость для калия и его поток из клетки инактивирует натриевую проницаемость и приводит к восстановлению заряда на мембране. Возникает фаза реполяризации.
Существенным фактором является натрий-калиевый насос, который выводит из клетки 3 иона натрия в обмен на 2 иона калия, вводимые в клетку. Его работа зависит от метаболизма клетки, в частности, от ее энергоснабжения. При этом расходуется 1 молекула АТФ (рис.6).

Рисунок 6. Механизм работы натрий-калиевого насоса.
ПД состоит из пикового потенциала, который образуется фазой деполяризации, реверсии и реполяризации, и следовых потенциалов (рис.2).
Следовые потенциалы:
Отрицательный (следовая деполяризация);
Положительный (следовая гиперполяризация).
Причиной следовых потенциалов являются дальнейшие изменения соотношения между входом натрия в клетку и выходом калия из нее. При следовой деполяризации отмечается остаточный ток натрия в клетку при одновременном снижении калиевого тока. При следовой гиперполяризации – остаточное усиление тока калия из клетки при одновременной активации натрий-калиевого насоса.
ПД характеризуется:
1. изменяющимся характером;
2. кратковременностью – несколько мсек;
3. зарядом мембраны, снаружи – «-», внутри – «+».
При действии веществ, блокирующих натриевые каналы, ПД не генерируется, т.к. в норме деполяризация мембраны обусловлена повышение ее натриевой проницаемости. При увеличении силы раздражителя выше порога амплитуда ПД не изменяется, т.к. не изменяется число активированных натриевых каналов, которые максимально раскрываются при пороговом раздражении.
Условия, необходимые для возникновения возбуждения (законы раздражения).
Возбудимость тканей различна. Чтобы вызвать возбуждение, раздражитель должен обладать:
1. Достаточной силой – закон порога.
2. Крутизной (градиентом) нарастания этой силы – закон аккомодации.
3. Временем действия – закон силы-времени.
1. Закон силы.
Мерой возбудимости является порог раздражения – минимальная сила раздражителя, способная вызвать возбуждение. Все раздражители можно разделить на подпороговые, пороговые и сверхпороговые. По биологическому значению раздражители делят на адекватные (действующие на ткань в естественных условиях, к ним она приспособлена в процессе эволюции) и неадекватные. В физиологических экспериментах в качестве раздражителя чаще всего используется электрический ток, т.к. он вызывает обратимые изменения, легко дозируется по силе и длительности, по своей природе близок к электрическим процессам, протекающим в живых организмах.
В 1870 г. Боудич в эксперименте на мышце сердца путем нанесения на неё одиночных пороговых раздражений регистрировал ответную реакцию - установил, что на подпороговое раздражение реакции не было, при пороговой силе и сверхпороговой амплитуда ответной реакции была одинаковой. На основании этого он предложил закон «Всё или ничего».
После введения в экспериментальные исследования микроэлектронной техники было установлено, что на подпороговое раздражение в ткани возникает ответная реакция. Если сила стимула меньше 50% пороговой величины, то под полюсами электродов происходит пассивная деполяризация без изменения проницаемости мембраны для ионов (электротонические изменения). Если сила стимула меньше пороговой величины, но больше 50% от нее, то в ткани возникает локальный ответ, который сопровождается деполяризацией мембраны в области нанесения раздражения и не распространяется на всю ткань, возбудимость тканей в этом участке повышена. Локальный ответ подчиняется закону силовых отношений, т.е. чем больше сила подпорогового стимула, тем больше амплитуда локального ответа. Проницаемость мембраны клетки в этом участке повышается для ионов натрия. При нанесении порогового стимула возникает ПД, амплитуда которого не изменяется, если величина стимула будет превышать пороговую, т.е. отвечает закону «Все или ничего», но на сверхпороговые стимулы длительность ПД будет меньше за счет укорочения продолжительности локального ответа.
Момент перехода локального ответа в ПД называется критическим уровнем деполяризации (КУД), а сдвиг заряда мембраны с мембранного потенциала до КУД, называется пороговым потенциалом, он наряду с порогом раздражения характеризует возбудимость ткани.
Изменение возбудимости тканей при возбуждении.
При возбуждении возбудимость тканей претерпевает определенные изменения в зависимости от фаз ПД (рис.7):
I – супернормальная возбудимость (первичная) соответствует локальному ответу, при этом два подпороговых стимула, нанесенных с интервалом времени, короче длительности локального ответа могут суммироваться и вызывать ПД;
II – абсолютная рефрактерность – соответствует регенеративной деполяризации и реверсии, при этом ткань становится абсолютно невозбудимой и не отвечает на самые сильные раздражители;
III – относительная рефрактерная фаза, соответствует реполяризации, при этом возбудимость ткани постепенно восстанавливается и сверхпороговый стимул, нанесенный в этот период может генерировать ПД;
IV – супернормальная возбудимость (вторичная или экзальфационная фаза) - следовой деполяризации, ткань становится более возбудимой, чем в исходном состоянии и даже подпороговый стимул способен вызвать ПД;
V – субнормальная возбудимость – следовой гиперполяризации, возбудимость ткани несколько снижена.

Рисунок 7. Изменение возбудимости мембраны при развитии потенциала действия.
2. Закон градиента раздражения (Дюбуа Реймон).
Чем больше градиент раздражения, тем больше (до известных пределов) реакция живого образования.
За время действия медленно нарастающего стимула наступает приспособление ткани – аккомодация. Она связана с тем, что при возбуждении проницаемость для ионов натрия увеличивается на короткий промежуток времени, если в течение его раздражитель не достигает пороговой величины, то увеличивающаяся проницаемость для ионов калия инактивирует натриевую проницаемость и возбуждение не наступает. При этом происходит также сдвиг КУД с увеличением порогового потенциала.
3. Закон силы-времени (Лапик).
Пороговая величина любого раздражителя находится в обратной зависимости от времени его действия, которая характеризуется математической кривой – гиперболой. Характер кривой свидетельствует о том, что подпороговые стимулы (меньше 1 реобазы) не вызовут возбуждение как долго бы они не действовали, в то же время очень сильный кратковременный стимул, длительность которого меньше полезного времени, также не вызовет возбуждение.
Сила постоянного тока, которая, действуя неопределенное время, вызывает возбуждение, называется реобазой.
Время, в течение которого ток в 1 реобазу вызывает возбуждение – полезное время.
Минимальное время, в течение которого ток силой в 2 реобазы вызовет возбуждение, называется хронаксией. Исследование этого показателя используется в неврологической и травматологической практике для изучения динамики восстановления в нервной или мышечной ткани после травмы.
Список использованной литературы
- Нормальна фізіологія /Під ред. В.І. Філімонова. – К. – Здоров’я, 1994. – С. 5 - 37.
- Физиология человека /Под ред. Г. И. Косицкого. – М., Медицина, 1985. – С. 19 – 84.
- Посібник з нормальної фізіології /Під ред. В.Г. Шевчука. – К., Здоров’я, 1995. – С. 6 - 36.
- Руководство к практическим занятиям по физиологии /Под ред. Г. И. Косицкого. – М., Медицина, 1988. – С. 72 - 94.
- Нормальная физиология /Под ред. В. И. Филимонова. - Запорожье, 1995. – С. 74-72.
- Физиология человека. Т.1 /Под ред. Р. Шмидта и Г. Тевса. – М., Мир, - 1996. - С. 9-87.
- Физиология человека. Т.1 / Под ред. В.М. Покровского. – М., Медицина, 1998. – С. 27-97.
- Общий курс физиологии человека и животных. Т.1. /Под ред. А.Д. Ноздрачева – М., Высшая школа, 1991.- С.36-116.
- Физиология человека. /Под ред. В.М. Смирнова – М., Медицина, 2002. – С. 45-61, 82-94.
- Фізіологія людини. Вільям Ф. Ганонг. – БаК, Львів, 2002. – С. 6 – 69, 74-76.
И потенциал действия большинства нейронов. Тем не менее, в основе любого потенциала действия лежат следующие явления:
- Мембрана живой клетки поляризована - её внутренняя поверхность заряжена отрицательно по отношению к внешней благодаря тому, что в растворе возле её внешней поверхности находится бо́льшее количество положительно заряженных частиц (катионов), а возле внутренней поверхности - бо́льшее количество отрицательно заряженных частиц (анионов).
- Мембрана обладает избирательной проницаемостью - её проницаемость для различных частиц (атомов или молекул) зависит от их размеров, электрического заряда и химических свойств.
- Мембрана возбудимой клетки способна быстро менять свою проницаемостъ для определённого вида катионов, вызывая переход положительного заряда с внешней стороны на внутреннюю (Рис.1 ).
Первые два свойства характерны для всех живых клеток. Третье же является особенностью клеток возбудимых тканей и причиной, по которой их мембраны способны генерировать и проводить потенциалы действия.
Фазы потенциала действия
- Предспайк - процесс медленной деполяризации мембраны до критического уровня деполяризации (местное возбуждение, локальный ответ).
- Пиковый потенциал, или спайк, состоящий из восходящей части (деполяризация мембраны) и нисходящей части (реполяризация мембраны).
- Отрицательный следовой потенциал - от критического уровня деполяризации до исходного уровня поляризации мембраны (следовая деполяризация).
- Положительный следовой потенциал - увеличение мембранного потенциала и постепенное возвращение его к исходной величине (следовая гиперполяризация).
Общие положения
Поляризация мембраны живой клетки обусловлена отличием ионного состава с её внутренней и наружной стороны. Когда клетка находится в спокойном (невозбуждённом) состоянии, ионы по разные стороны мембраны создают относительно стабильную разность потенциалов, называемую потенциалом покоя . Если ввести внутрь живой клетки электрод и измерить мембранный потенциал покоя, он будет иметь отрицательное значение (порядка −70 - −90 мВ). Это объясняется тем, что суммарный заряд на внутренней стороне мембраны существенно меньше, чем на внешней, хотя с обеих сторон содержатся и катионы , и анионы . Снаружи - на порядок больше ионов натрия , кальция и хлора , внутри - ионов калия и отрицательно заряженных белковых молекул, аминокислот, органических кислот, фосфатов , сульфатов . Надо понимать, что речь идёт именно о заряде поверхности мембраны - в целом среда и внутри, и снаружи клетки заряжена нейтрально.
Потенциал мембраны может изменяться под действием различных стимулов. Искусственным стимулом может служить электрический ток , подаваемый на внешнюю или внутреннюю сторону мембраны через электрод. В естественных условиях стимулом часто служит химический сигнал от соседних клеток, поступающий через синапс или путём диффузной передачи через межклеточную среду. Смещение мембранного потенциала может происходить в отрицательную (гиперполяризация ) или положительную (деполяризация ) сторону.
В нервной ткани потенциал действия, как правило, возникает при деполяризации - если деполяризация мембраны нейрона достигает некоторого порогового уровня или превышает его, клетка возбуждается, и от её тела к аксонам и дендритам распространяется волна электрического сигнала. (В реальных условиях на теле нейрона обычно возникают постсинаптические потенциалы, которые сильно отличаются от потенциала действия по своей природе - например, они не подчиняются принципу «всё или ничего». Эти потенциалы преобразуются в потенциал действия на особом участке мембраны - аксонном холмике, так что потенциал действия не распространяется на дендриты).
Рис. 3. Простейшая схема, демонстрирующая мембрану с двумя натриевыми каналами в открытом и закрытом состоянии, соответственноЭто обусловлено тем, что на мембране клетки находятся ионные каналы - белковые молекулы, образующие в мембране поры, через которые ионы могут проходить с внутренней стороны мембраны на наружную и наоборот. Большинство каналов ионоспецифичны - натриевый канал пропускает практически только ионы натрия и не пропускает другие (это явление называют селективностью). Мембрана клеток возбудимых тканей (нервной и мышечной) содержит большое количество потенциал-зависимых ионных каналов, способных быстро реагировать на смещение мембранного потенциала. Деполяризация мембраны в первую очередь вызывает открытие потенциал-зависимых натриевых каналов. Когда одновременно открывается достаточно много натриевых каналов, положительно заряженные ионы натрия устремляются через них на внутреннюю сторону мембраны. Движущая сила в данном случае обеспечивается градиентом концентрации (с внешней стороны мембраны находится намного больше положительно заряженных ионов натрия, чем внутри клетки) и отрицательным зарядом внутренней стороны мембраны (см. Рис. 2). Поток ионов натрия вызывает ещё бо́льшее и очень быстрое изменение мембранного потенциала, которое и называют потенциалом действия (в специальной литературе обозначается ПД).
Согласно закону «всё-или-ничего» мембрана клетки возбудимой ткани либо не отвечает на стимул совсем, либо отвечает с максимально возможной для неё на данный момент силой. То есть, если стимул слишком слаб и порог не достигнут, потенциал действия не возникает совсем; в то же время, пороговый стимул вызовет потенциал действия такой же амплитуды , как и стимул, превышающий пороговый. Это отнюдь не означает, что амплитуда потенциала действия всегда одинакова - один и тот же участок мембраны, находясь в разных состояниях, может генерировать потенциалы действия разной амплитуды.
После возбуждения нейрон на некоторое время оказывается в состоянии абсолютной рефрактерности , когда никакие сигналы не могут его возбудить снова, затем входит в фазу относительной рефрактерности , когда его могут возбудить исключительно сильные сигналы (при этом амплитуда ПД будет ниже, чем обычно). Рефрактерный период возникает из-за инактивации быстрого натриевого тока, то есть инактивации натриевых каналов (см. ниже).
Распространение потенциала действия
Распространение потенциала действия по немиелинизированным волокнам
По ходу ПД каналы переходят из состояния в состояние: у Na + каналов основных состояний три - закрытое, открытое и инактивированное (в реальности дело сложнее, но этих трёх достаточно для описания), у K + каналов два - закрытое и открытое.
Поведение каналов, участвующих в формировании ПД, описывается через проводимость и высчиляется через коэффициенты переноса (трансфера).
Коэффициенты переноса были выведены Ходжкиным и Хаксли.
Проводимость для калия G K на единицу площади
| , |
| где: |
| - коэффициент трансфера из закрытого в открытое состояние для K+ каналов ; |
| - коэффициент трансфера из открытого в закрытое состояние для K+ каналов ; |
| - фракция К+ каналов в открытом состоянии; |
| - фракция К+ каналов в закрытом состоянии |
Проводимость для натрия G Na на единицу площади
рассчитывается сложнее, поскольку, как уже было сказано, у потенциал-зависимых Na+ каналов, помимо закрытого/открытого состояний, переход между которыми описывается параметром , есть ещё инактивированное/не-инактивированное состояния, переход между которыми описывается через параметр
| , | , |
| где: | где: |
| - коэффициент трансфера из закрытого в открытое состояние для Na+ каналов ; | - коэффициент трансфера из инактивированного в не-инактивированное состояние для Na+ каналов ; |
| - коэффициент трансфера из открытого в закрытое состояние для Na+ каналов ; | - коэффициент трансфера из не-инактивированного в инактивированное состояние для Na+ каналов ; |
| - фракция Na+ каналов в открытом состоянии; | - фракция Na+ каналов в не-инактивированном состоянии; |
| - фракция Na+ каналов в закрытом состоянии | - фракция Na+ каналов в инактивированном состоянии. |
См. также
Литература
Wikimedia Foundation . 2010 .
2.2. Потенциал действия: фазы потенциала действия, механизм возникновения. Восстановительный период. Явление аккомодации возбудимой ткани.
Потенциал действия . Если участок нервного или мышечного волокна подвергнуть действию достаточно сильного раздражителя (например, толчка электрического тока), в этом участке возникает возбуждение, одним из наиболее важных проявлений которого служит быстрое колебание МП, называемое потенциалом действия (ПД)
Причиной возникновения ПД является изменение ионной проницаемости мембраны. В состоянии покоя, как уже говорилось, проницаемость мембраны для К + превышает натриевую проницаемость. Вследствие этого поток положительно заряженных ионов из протоплазмы наружу превышает противоположный поток Na + . Поэтому мембрана в покое снаружи заряжена положительно.
При действии на клетку раздражителя проницаемость мембраны для ионов Na+ резко повышается, и в конечном итоге становится примерно в 20 раз больше проницаемости для К + . Поэтому поток ионов Na + в клетку начинает значительно превышать направленный наружу поток К + . Ток Na + достигает величины +150 мв. Одновременно несколько уменьшается выход К + из клетки. Все это приводит к извращению (реверсии) МП, и наружная поверхность мембраны становится заряженной электро отрицательно по отношению к внутренней поверхности. Указанный сдвиг и регистрируется в виде восходящей ветви пика ПД (фаза деполяризации).
При внутриклеточном отведении можно обнаружить, что поверхность возбужденного участка на очень короткий интервал, измеряемый тысячными долями секунды, становится заряженным электроотрицательно по отношению к соседнему, покоящемуся участку, т.е. при возбуждении происходит т.н. "перезарядка мембраны". Точные измерения показали, что амплитуда ПД на 30-50 мв превышает величину МП. Причина этого состоит в том, что при возбуждении происходит не просто исчезновение ПП, а возникает разность потенциалов обратного знака, в результате чего наружная поверхность мембраны становится заряженной отрицательно по отношению у ее внутренней стороне.
Потенциал действия протекает фазно. Временной ход потенциала действия включает четыре последовательных этапа: локальный ответ, деполяризацию, реполяризацию и следовые потенциалы (рис. 2). В ПД принято различать его пик (т.н. спайк - spike) и следовые потенциалы. Пик ПД имеет восходящую и нисходящую фазы. Перед восходящей фазой регистрируется более или менее выраженный т.н. местный потенциал, или локальный ответ. Поскольку во время восходящей фазы исчезает исходная поляризация мембраны, ее называют фазой деполяризации; соответственно нисходящую фазу, в течение которой поляризация мембраны возвращается к исходному уровню, называется фазой реполяризации. Продолжительность пика ПД в нервных и скелетных мышечных волокнах варьирует в пределах 0,4-5,0 мсек. При этом фаза реполяризации всегда продолжительнее.
Рис. 2. Фазы и временной ход потенциала действия.
Кроме пика, в ПД различают два следовых потенциала - следовую деполяризацию (следовой отрица-тельный потенциал) и следовую гиперполяризацию (следовой положи-тельный потенциал. Амплитуда этих потенциалов не превышает нескольких милливольт, а длительность варьирует от нескольких десятков до сотен миллисекунд. Следовые потенциалы связаны с восстановительными процессами, развивающимися в мышцах и нерве после окончания возбуждения.
Промежуток времени, в течение которого сохраняется активное состояние в виде ПД, неодинаков в разных возбудимых структурах. В нейронах он составляет около 1 мс, в волокнах скелетных мышц – 10 мс, в миокарде достигает 200–250 мс.
Левое крыло графической записи ПД, отражающее изменение потенциала в электроположительную сторону называется деполяризацией. Область электроположительности носит название овершута, правое крыло ПД, свидетельствующее о восстановлении исходного поляризованного состояния мембраны принято называть реполяризацией. Часто, но не всегда возвращение ПД к исходному уровню в состоянии покоя происходит с наличием фаз в форме так называемых следовых потенциалов. Следовые потенциалы неодинаковы в мышцах и нервах. В волокнах скелетных мышц фаза реполяризации очень замедлена. Примерно через 1 мс после начала ПД наблюдается отчетливый перегиб крыла реполяризации – это следовая деполяризация. В нейроне чаще всего кривая реполяризации быстро пересекает уровень МПП и на некоторое время потенциал мембраны становится более электроотрицательным, чем МП. Это явление называют следовой гиперполяризацией.
Повышение проницаемости мембраны для ионов Na + продолжается в нервных клетках очень короткое время. Связано оно с кратковременным открытием т.н. Na + -каналов (точнее, заслонок М в этих каналах), которое затем сменяется срочным закрытием Na+-пор с помощью т.н. Н-ворот. Этот процесс называется натриевой инактивацией. В результате поток Na в клетку прекращается.
Наличие специальных Na- и К- каналов и сложного механизма запирания и открытия ворот изучено биофизиками достаточно хорошо. Показано, что существуют избирательные механизмы, регулирующие те или иные каналы. Например, яд тетродотоксин блокирует только Na-поры, а тетраэтиламмоний - только К-поры. Показано, что у некоторых клеток возникновение возбуждения связано в изменением проницаемости мембраны для Са ++ , в других - для Mg + . Исследования механизмов изменения проницаемости мембран продолжаются.
В результате Na-инактивации и одновременного увеличения К- проницаемости происходит усиленный выход положительных ионов К+ из протоплазмы во внешний раствор. В итоге этих двух процессов происходит восстановление поляризованного состояния мембраны (реполяризация) , и наружная ее поверхность вновь приобретает положительный заряд. В дальнейшем происходят процессы восстановления нормального ионного состава клетки и необходимого градиента концентрации ионов за счет активизации деятельности Na-К-насоса. В результате повышения проводимости резко возрастает поток катионов Na + , поэтому отрицательный заряд в клетке вблизи внутренней стороны поверхности мембраны также резко уменьшается вплоть до преобладания положительных зарядов. В результате происходит изменение знака потенциала, достигающего +30 мВ. После этого проводимость мембраны дляNa + также резко снижается.
Для нормального протекания ПД играет существенную роль и изменение проводимости мембраны для K + , которая начинает возрастать позже возрастания проводимости дляNa + . Увеличение относительно медленного выходаK + из клетки в фазу снижения проводимости дляNa + вызывает реполяризацию мембраны.
Таким образом, в живой клетке существуют два различных типа движения ионов через мембрану. Один из них осуществляется по градиенту концентрации ионов и не требует затраты энергии, поэтому его называют пассивным транспортом. Он ответственен за возникновение МП и ПД и ведет в конечном итоге к выравниванию концентраций ионов по обе стороны клеточной мембраны. Второй тип движения ионов через мембрану, осуществляющийся против концентрационного градиента, состоит в "выкачивании" ионов Na+ из протоплазмы и "нагнетании" ионов К+ внутрь клетки. Этот тип ионного транспорта возможет лишь при условии затраты энергии - это активный транспорт. Он является результатом работы специальных ферментных систем (т.н. насосов), и благодаря ему восстанавливается исходная разность концентраций, необходимая для поддержания МП.
Условия возникновения возбуждения . Для возникновения ПД необходимо, чтобы под влиянием какого-либо раздражителя произошло повышение ионной проницаемости мембраны возбудимой клетки. Однако, возбуждение возможно лишь при условии, если действующий на мембрану агент имеет некоторую минимальную (пороговую) величину, способную изменить мембранный потенциал (МП, или Ео) до некоторого критического уровня (Ек, критический уровень деполяризации). Стимулы, сила которых ниже пороговой величины, называются подпороговыми, выше - надпороговыми. Показано, что пороговая сила, необходимая для возникновения возбуждения при внутриклеточном микроэлектроде равна 10 -7 - 10-9 А.
Таким образом, главным условием для возникновения ПД является следующее: мембранный потенциал должен стать равным или меньше критического уровня деполяризации (Ео <= Eк)
Инактивация Na+-системы. Na+-системой обозначают механизм, позволяющий в течение нескольких долей миллисекунды многократно (до 20 раз) увеличить проводимость клеточной мембраны для Na+. Достигнув пикового значения, примерно через 0,5 мс проводимость мембраны для Na+ начинает снижаться. Быстрое снижение проводимости для Na+ называют инактивацией Na+-системы. В основе инактивации Na+-системы лежит переход в инактивационное состояние потенциалзависимых Na+-каналов. Поэтому скорость и степень снижения проводимости потенциалзависимы. Это означает, что чем больше отличается потенциал мембраны от мембранного потенциала покоя в сторону электроположительности, тем сильнее инактивирована Na+-система. Поэтому деполяризация мембраны вызывает снижение тока Na+ внутрь клетки. С одной стороны, это свидетельствует о том, что усиление тока Na+ само себе служит причиной его быстрого последующего снижения и начала развития реполяризации. С другой стороны, это означает, что если исходный потенциал клетки выше потенциала покоя на 20–30 мВ, то Na+-система полностью инактивирована и никакая последующая деполяризация уже не может активировать ее, т.е. вызвать резкое увеличение проводимости для Na+ и генерацию ПД.

